

亜高山の落葉樹林下や、草原に生える多年草です。 茎は高さ10〜30cmほど。 本州中部以北と北海道に分布します。 葉は2個あり、互生ですが接近してほぼ対生し、広楕円形。 長さは10〜15cm、幅は4〜10cm(*3)。 写真#1では、花をつけた株の手前に、まだ花茎を伸ばしていない株の葉が見えます。
竹の根を細くしたような地下茎が、地中浅く所々で分岐しながら縦横に走り、長いものでは1m以上にもなるそうです(*4)。 ここから根を出し、栄養繁殖も行います(*5)。

6〜7月に葉の間から緑色で毛が密生した茎を伸ばし、茎頂に淡黄緑色の花を横向きに1個だけ咲かせます。 花は淡黄緑色で紫褐色の斑があり、長さは4cm、幅は3cmほど(*1,*4)。 変わった形をしていて、ウツボカズラの食中袋を連想しました。
この花の自生地については、大雑把な場所は知っていたのですが、そこにたどり着くには、山登りが超苦手な私たちでは、かなりの時間を要すると思われました。 仮にすごく頑張ってたどり着けたとしても、下山する体力を使い果たしてしまっている可能性が高いと思われるほどの場所です。 このため、一生見ることは叶わないだろうと諦めていたのですが... ある方から別の自生地を教えていただき、念願叶って見ることができました。 情報をご提供下さった方に本当に感謝いたします。

花の上に覆いかぶさるようになっている白色の部分は、背萼片です。 広卵形で、長さは2〜2.5cmほど(*3)。 背萼片の内面は緑褐色ですが、外面は縁の部分のみ褐色で、残りは全面白色で反射率が高く、明るい陽光の下では白飛びしそうで、撮影には気を使いました。
この自生地はすぐ間近に樹木がない、明るい草地でした。 背萼片は蕊柱や唇弁に覆いかぶさるようになっていますが、雨からこれらの大切な器官を護ると同時に、直射日光による花の温度上昇を軽減する役割もあると推測します。 白色の外面で太陽光を反射し、それでも通過してきた光を、内面の褐色の色素で遮断するのでしょう。
次に花の詳細を見ていきますが、変わった形の花なので、細かく図示しました。

写真#4は、本種の花をほぼ正面から見たものです。 大きな背萼片の下には、左右に開いた側花弁があります。 側花弁は全体として白色で、緑褐色の班があり、線状長楕円形で先端はへら状に広がり円頭、基部の内面に密毛があります。 ほぼ白色で左右に大きく開いているので目立ち、「花はココですよ!」と広告塔の役割もありそうです。
蕊柱(ずいちゅう)については、写真#9以降で詳しく観察します。
唇弁は最も目を引く部分で、本種は底の丸いコップのような形状です。 図鑑によってはこれを「壺状」や「袋状」と表現しています(アツモリソウやクマガイソウの花は「袋状」が適切と思います)。 開口部は広く、つぼまってなく、アツモリソウやクマガイソウなど他のアツモリソウ属の植物と大きく異なる点です。
唇弁は淡黄色と淡褐色のまだら模様になっています。 筆者はクマガイソウの観察において唇弁に光を通しやすい部分が存在することに気付き、これは唇弁内部に入った昆虫を誘導するための仕組みであるという「半透明点唇弁上部多分布説」を展開しましたが、それを思い起こさせる模様です。
しかし本種の唇弁は開口部が広く、「返し」もないので、唇弁内に落ちた昆虫は容易に開口部から逃げられるようにも思えます。 本種において唇弁のまだら模様がどのような意味を持つのかは、よくわかりません。

#5は本種の花の側面です。 側面から見ると、唇弁の開口部が単純な丸い形ではなく、後方から前方にかけて緩やかな曲線を描いて下がり「受け口」のようになっていることがわかります。 何を「受ける」のでしょうか? それは花粉を運んでくれる送粉者の昆虫に違いないでしょう。 送粉者がコップ状の唇弁の中に落ちるように意図された形状に見えます。
花柄子房にはねじれはなく、近縁種のアツモリソウやクマガイソウと同様「ストレート・唇弁下側タイプ」です。 側面から見ると、苞や合萼片も見えてきます。 これらを次の写真で詳しく見ます。

写真#6は、花の背面を後方斜め上から見たものです。 苞、背萼片、花茎にびっしりと腺毛が生えている様子がわかります。 背萼片が雨や太陽光から花を護るように覆っていることは上で述べましたが、苞(苞葉)はさらに背萼片を護るかのように覆いかぶさっているのです。
苞には、花がまだつぼみの時に包んで護る役割があります。 他の多くの種類の植物では、開花後は役割を終え、まるで飾りのように花の下についているものです(開花後も重要な役割を果たす種もあります)。 しかし本種では開花後も太陽光の熱や紫外線から花を護るが如く、最後の最後まで一番上で花を護り続けているかのように見えます。
ところでラン科植物の花には、必ず3個の内花被片と3個の外花被片があります。 内花被片は、2個の側花弁と1個の唇弁。 外花被片は、1個の背萼片と2個の側萼片です。 本種の側萼片は? 実は、アツモリソウ属は、2個の側萼片が合着し「合萼片(ごうがくへん)」と呼ばれる1個の萼片になっているのです。 次の写真でもう少し詳しく見ましょう。


他のラン科植物では2個ある側萼片が、本種が属するアツモリソウ属では1個に合着し、合萼片となって唇弁の真後ろにあります(#7)。 #8は#7の長方形枠部の拡大です。 合萼片の先端部をよく見ると、わずかに2裂していることがわかります。
これはかつては2個の側萼片であった名残だと思います。
なぜアツモリソウ属において側萼片が合萼片となったのか、それを明確に示した文献はまだ見たことがなく、不明です。 しかし1個にまとめられ花の背後に配置された状態を見ると、アツモリソウ属においては側萼片が邪魔なものになったのではないかと推測したくなります。 邪魔になった原因は、アツモリソウ属の特殊な形状の唇弁と、送粉者の誘導方法に関係があると思います。
すなわちアツモリソウ属は、特殊な袋状やコップ状の唇弁をつくり、内部に入った昆虫を「正の走光性」を利用して柱頭・花粉がある出口方向へ導く必要があるのですが、側花弁が光を遮るため、始めはそれがうまくいかなかったと考えられます。 正の走光性を利用するためには、唇弁の特に側面から十分に光が当たる必要があるので、光を遮る2つの側萼片を1つにまとめて、花の後方に追いやったのではないでしょうか。 突飛な考えと思われるかも知れませんが、このように考えると合萼片となった理由が説明できると思います(あくまでも筆者の私見ですが)。
次は、いよいよ花の中心部に迫ります。

#9は花の中心部を正面から見たものです。 仮雄しべは縦長のチョウの翅のようになっていて、両側が手前側に折れ曲がっているようになっています(#10でよくわかります)。 両側に小さく突き出しているものは雄しべですが、仮雄しべの後方にあります。
仮雄しべは、その位置・大きさ・色から、送粉者となる昆虫の着地地点となると思い割れます。 しかしその表面をよく見ると、縦方向に細かな溝があることがわかります。 これはあたかも昆虫が着地したときに滑りやすくしているように見えます。昆虫が仮雄しべに降りたときに足を滑らせたらどうなるか? もちろん、唇弁の中に落ちるでしょう。 それを狙っていると思える形状なのです。 なんとも巧妙なものです。
一般にラン科の植物は3個の雄しべと1個の雌しべを持つ祖先から進化したとされていますが、普通のラン科植物ではそれらが融合して蕊柱を作ります。 しかしアツモリソウ亜科の植物では、それらの融合が不完全なのです。 また普通のランでは3個の雄しべの内で外側の1個が残って髄柱の先端に位置し、あとの2個は退化しているのに対して、アツモリソウ亜科では雄しべのうち、内側の2個が残り、雌しべの基部の両側に位置します。 そして先端の上側には仮雄蘂があり、その下面に柱頭があるという構造なのです(*8)。
この雄しべ・仮雄しべの特殊な構造は、コップ状・袋状の唇弁に送粉者を誘い込み、脱出するときに花粉をくっつけるという、アツモリソウ亜科植物の花粉媒介の方法と密接な関係があると思えます。
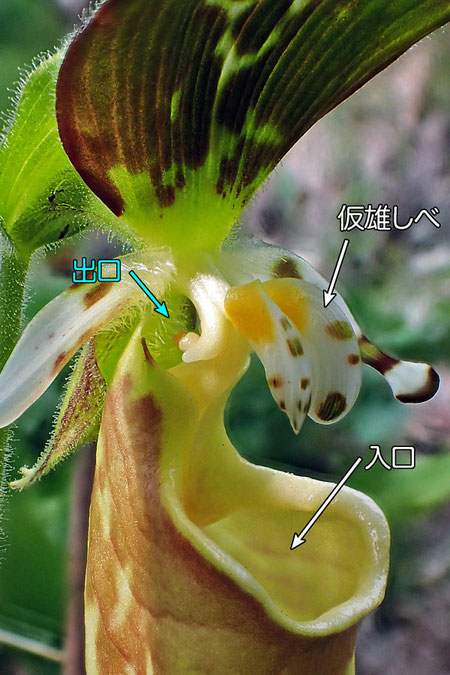
大きな仮雄しべは上部が黄色く、あるよく目立ち、昆虫に「蜜がありそうだ」と思わせるかも知れません。 仮雄しべはある程度大きいので、飛んできた昆虫はここに着地を試みます。 しかし着地してみると、つかまりにくい形状のため、「入口」である唇弁の開口部から中に落ちてしまいます。 開口部の縁につかまりかけますが、滑りやすい形状なので、こらえきれずに中に落ちます。
唇弁の中にあると期待していた蜜は.... ありません。 本種は蜜がありそうな姿をしていながら、蜜持っていないのです。 仕方なく、昆虫は唇弁の奥に生えた毛を足がかりに「出口」を目指して登っていきます。
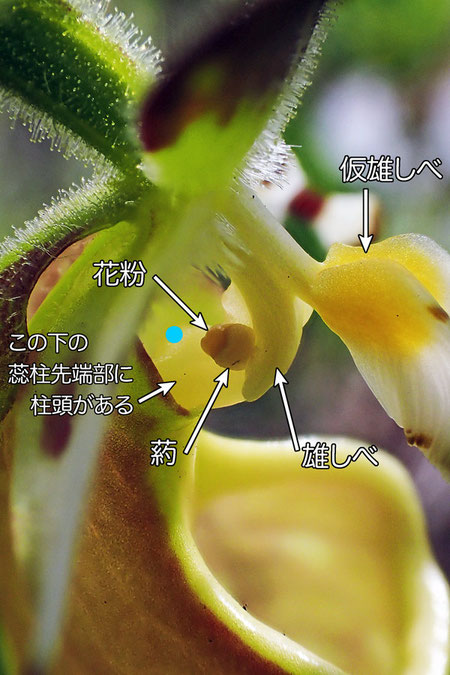
1個の仮雄しべ、2個の雄しべと葯、1個の雌しべ(とその柱頭)、これらをまとめて蕊柱といいます。 #11では雌しべはちょっと見づらく、その先端の柱頭は花の中に隠れて、残念ながら見えません。
唇弁の奥の壁を登って来た昆虫は、出口付近が狭いので、ちょっと苦労します。 もしこの昆虫がすでに他のキバナノアツモリソウの花を訪れていたなら、背中にその花粉をつけている可能性が高い。 そして出口手前にある柱頭のそばをすり抜けるときに、背中の花粉が柱頭に付着して受粉します。
さらに進むと、間もなく出口です。 #11では、青い●印あたりです。 ここも狭く、やっとこさすり抜けますが、どうしても雄しべの葯の先についた「花粉」に背中をこすりつけないと通れません。
多くのラン科植物の花粉は、非常に細かく(0.1〜0.5mm程度)かつ多数(数万〜数百万個)の花粉が、蠟で固められたような「花粉塊(かふんかい)」をつくります(*7)。 しかしアツモリソウ亜科の植物では、花粉塊をつくらず、花粉が柔らかい粘液質でまとめられていることが特徴です。 #11で「花粉」と指し示した半球状の部分には、とんでもなくたくさんの微細な花粉が含まれているのです。
昆虫が「花粉」に背中をこすりつけて脱出するとき、背中には多数の花粉が含まれた粘着質のものが付着します。 この状態でまた別の花を訪れると、同じような受粉が起こるのです。 但し蜜がないことを昆虫が学習すると、再び本種に訪花する可能性は低くなります。 事実、本種へ昆虫が訪れることはあまり多くないようです。
〜 最後に 〜
元々数が少ない植物のようです。 アツモリソウほどではないにしろ、盗掘が絶えず、減少し続けています。 環境省が絶滅危惧種Ⅱ類に指定している希少植物です。
もし幸運にも出逢えたら、そっと見守ってあげて下さい。 また思わぬ情報拡散により盗掘を誘発させないためにも、自生地情報の取扱は要注意だと思います。

< 引用・参考文献、及び外部サイト(順不同、敬称略) >
*1 牧野 新日本植物圖鑑 北隆館 1961年6月30日初版発行 p.877
山と渓谷社 2013年3月30日 初版第1刷 p.130
*3 日本の野生植物 草本 1 単子葉類 平凡社 1982年1月20日 初版第1刷 p.191
*4 日本の野生ラン 主婦と生活社 1996年 43刷 発行 p.10
*5 ラン科アツモリソウ属の生態〜最近の研究を中心に 丹波真一
北方山草会 会誌第26号(2009)
*6 日本産アツモリソウ属 Cypropedium(ラン科)の地理分布パターン 高橋英樹
日本植物分類学会 分類 Vol. 9 (2009) No. 2 p.143-157
*7 ランの王国 高橋英樹 2016 北海道大学出版会
*9 植物和名−学名インデックス YList キバナノアツモリソウ
※ 外部サイトは、それぞれの運営者の都合により、変更・削除されることがあります。
2018.05.10 掲載
他のラン科アツモリソウ属の植物










